Abstract / Introduction / Summary:
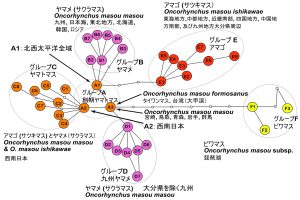
体側の赤点(あるいは朱点)の有無によって 陸封型のアマゴとヤマメは分類学的に区別されて きた(大島,1957).これらを含むサクラマス類 似種群は最近では 4 亜種(アマゴ Oncorhynchus masou ishikawae,ヤマメO. m. masou,ビワマスO. masou subsp.,およびタイワンマス O. m. formo- sanus)として分類されることが多い(Kottelat, 1996;McKay, 1998;細谷,2013).一方で,過去 には別種とする意見もあった(加藤,2002).し かしこれらを総合した遺伝学的解析と形態的解 析,及び動物命名規約に沿って十分学名を含めた 検討はされていない.琵琶湖のビワマスは陸封型 のアマゴの降湖タイプと長く考えられてきたが, 形態や生態も違うことからアマゴの降湖タイプか ら格上げして別種ではないかと考える研究者もい る(Fujioka and Fushiki, 1998;手賀ほか,1998; 加藤,2002;桑原・井口,2007;Kuwahara et al., 2012).Gwo (2008) は AFLP 解析の結果からサク ラマス類似種群はタイワンマスを含めて 4 亜種と した.Nakabo (2009) は計数・形態的特徴,過去 の遺伝学的解析結果による推定された分化年代 (Oohara and Okazaki, 1996),大分県玖珠盆地から 得られたビワマス類似化石の地質年代(Uyeno et al., 2000)からタイワンマスは最も原始的である と推定してタイワンマスを別種とした.これらの 経緯と研究経過を踏まえて,その後ビワマスはア マゴとは異なり,Nakabo (2009) の意見を踏襲し て亜種レベルとしている(細谷,2013).従って サクラマス類似種群は分類学的位置付けの合意は まだ得られていない状況であり,正しい学名を含 めて動物命名規約に沿って十分な分類学的検討も されていない.なお,アマゴとヤマメの降海型は それぞれサツキマスとサクラマスと呼ばれる. 日本では朱点が無いヤマメは関東以北の太平 洋岸と日本海側全域,国外では朝鮮半島とロシア のカムチャッカ半島南部,大分県の瀬戸内海の周 防灘と伊予灘に注ぐ河川を除く九州の河川に分布 するとされている.朱点が有るアマゴ は神奈川 県西部以西本州太平洋岸,四国,瀬戸内海に注ぐ 中国地方南部,四国北部,及び九州の大分県の一 部(山国川,駅館川,大分川,大野川等)の河川 に分布する(大島,1957).境界部分は大島ライ ンとして知られる.ビワマスは琵琶湖のみで知ら れ,タイワンマスは台湾の北部の大甲渓から知ら れる(Iwatsuki et al., 2019;Gwo, 2008). サクラマス類似種群における北西太平洋全体 や日本全国を網羅した地理的遺伝系統は十分明ら かになっていない.Yamamoto et al. (2019) は北海 道周辺を中心としたサクラマス(ヤマメ)と西南日本のサツキマス(アマゴ)分布域の代表的な地 点から得たアマゴ,更に琵琶湖のビワマスと台湾 のタイワンマスの 4 亜種から 4 亜種の遺伝学的解 析を行った(Yamamoto et al., 2019: fig. 4 参照). しかし 4 亜種は独立した明確なグループには分離 されておらず,ビワマスのみ他の亜種のハプロタ イプとやや離れた位置にグループを形成してい た.この理由は定かでないが,4 亜種を示唆する 解析領域の探索が必要かもしれない. 従って未だにサクラマス類似種群である 4 亜 種が,独立して 4 つの亜種グループが本当に存在 するのかも把握出来ていない.つまり,北西太平 洋に普通にみられ,日本人との関わりが大きいサ クラマス類似種群であるにも関わらず,形態や遺 伝学的な確かな証拠に基づく 4 亜種の存在も分類 学的に適格なのかどうかも不明瞭のままであると 言わねばならない. 戦後 75 年以上も経過したにも関わらず,サクラマス類似種群の地域的な在来判定の研究はなさ れたが,形態や遺伝学的解析から 4 亜種の独立性 の検討や,北西太平洋全体の地理的遺伝系統の特 徴も明確で無い.従って,増殖・保全事業におい て移植や放流時の科学的な 4 亜種の生物学的基礎 情報の不足から資源管理や保全対策が十分に立て られない状況である.またサクラマス類似種群の 4 亜種の正確な分化過程や,四国,紀伊半島など の地域に固有な系統が存在するのか,河川毎や支 流に固有な系統が実在するのかについても未だに 不明で,全体を見渡す明確な情報はなかった.こ れは全国のサクラマス類似種群の遺伝系統の基本 的な概観情報さえ無いので十分判断出来ない.つ まり資源・保全対策が科学的な視点に立った対策 が出来ないことを意味する状況である. そこで,上記の問題を解決するため今回遺伝 マーカーとして用いたのはミトコンドリア DNA チトクローム b 領域全域である.特に氷河期と間氷河期には影響を受けたであろうミトコンドリア DNA の呼吸代謝に関わるチトクローム b 領域に おいて情報を得ることとした.重要な進化速度の 違うミトコンドリア DNA の領域を混在させない ためにこの領域のみを調べた.また,地理的遺伝 情報が欠損しないようにチトクロームb は部分解 析ではなく全域解析(1141 bp)を行った. 次に北西太平洋におけるサクラマス類似種群 4 亜種の生息域全体を網羅するよう調査 ・ 解析し, まず地理的遺伝系統の概観情報を把握しようし た.その結果 6 つの遺伝グループと地理的遺伝系 統の概観の情報が得られた.幸運にも宮崎県串間 市の福島川では自然分布の可能性のあったヤマメ の固有な南限個体群の発見に繋がった.細胞のミ トコンドリア DNA の部品である ATGC(アデニ ン,チミン,グアニン,シトシン)の北西太平洋 の中で固有な遺伝配列であるハプロタイプ(配列 の組み合わせ)が福島川で確認されたのである. これは母系遺伝であり母親から子供に同じ配列が 遺伝する.この南限個体群は今回明らかにされた 九州のみで見られる特異なグループであるグルー プ D に入り,更にそのグループの特異な固有ハ プロタイプが 3 つも確認され,日本最南限個体群 として判断することが出来た(Iwatsuki et al., 2019). 全国の在来情報を提供してもらった内水面漁 協,県水試の研究者,および遊漁者から南限個体 群の証明と説明だけでなく,サクラマス類似種群 6 グループの生物学特性を日本語での紹介がほし いという要請を受けた.現時点でのサクラマス類 似種群の生物学的特性,日本列島の地史学的情報 や既存の最近の報告や未発表データ等を若干踏ま えて再考し,今後の検証・検討すべき内容等を述 べた.ご意見・ご批判をいただき,サクラマス類 似種群における今後の深い理解と更なる研究の発 展を期待する.